

あなたの「耳」はどこにある? コオロギ・バッタの耳の話

11月になり秋もますます深まってきました。
秋といえば様々な虫たちが鳴く季節 ―
マツムシのチンチロリン、スズムシのリーンリーン、エンマコオロギのコロコロリリリー…網戸から聞こえる様々な種の鳴き声に秋を感じますね。
私たちにとって秋の風物詩である虫の声も、鳴いている当人(虫?)からすればオスからメスへのラブコール。
後世へ子孫を残せるかどうかの真剣勝負です。

今回は秋の虫たちが属するバッタ目※の昆虫を例にお話しようと思います。
※目は生物を分類するときの階級です。大きい順に「界」「門」「綱」「目」(「亜目」)「科」「属」「種」に分けられます。
「キリギリス」と「バッタ」の違いは「触角の長さ」と「耳の位置」
10月に科学館でイベント「身近な昆虫採集体験」を開催しました。
子供たちに虫取り網、虫かごを貸し出して、自然観察園やサイエンスパークで昆虫を採集してもらいます。
採集した昆虫は館内サイエンスライブラリーの図鑑で種を調べます。


採集した昆虫は、夏の企画展「身近で気になる昆虫 Best 50」にランクインしたキマダラカメムシ、ツマグロヒョウモンをはじめ、クロウリハムシ、ヤマトシジミ、ルリシジミ、ウラナミシジミなど様々。
市街地でもたくさんの昆虫たちと出会うことができるのですね。
バッタ目に限定するとクビキリギス、ホシササキリ、ショウリョウバッタ、オンブバッタ、ショウリョウバッタモドキの計5種が獲られました。

参加者の皆さんは、バッタ目の種同定にかなり手こずっていた様子。体色が緑色や茶色などに偏り、どの個体も同じ種に見えてしまいますよね。
バッタ目同定の第一歩は、「キリギリス亜目」と「バッタ亜目」に分けることです。
両亜目を簡単に見分ける一番のポイントは「触角の長さ」。触角が体長よりも長ければキリギリス亜目、短ければバッタ亜目です。
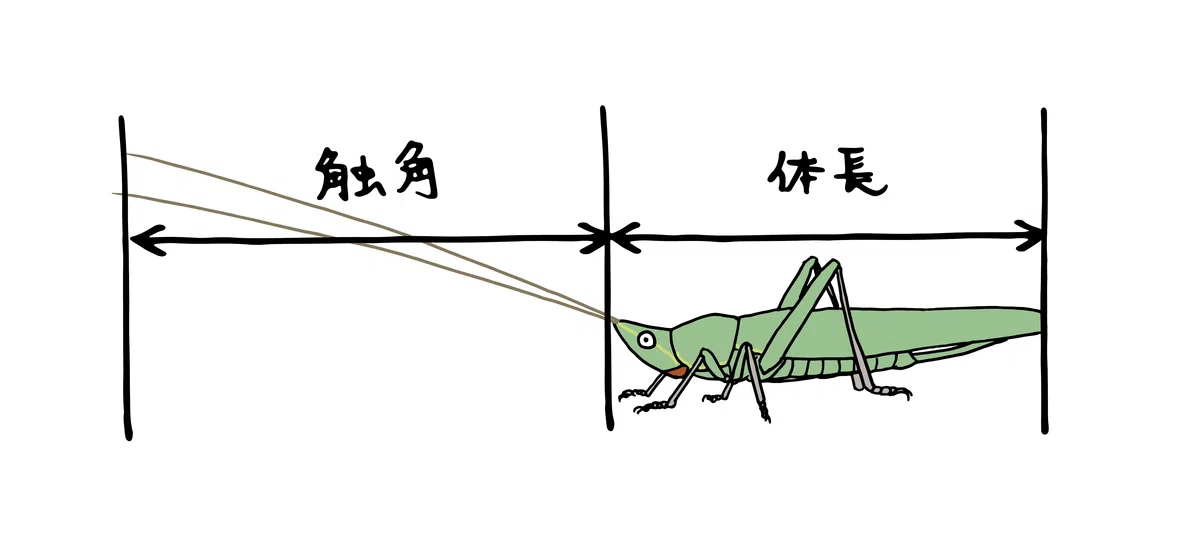
獲られた昆虫の触角の長さに着目してみると、
キリギリス亜目:クビキリギス、ホシササキリ
バッタ亜目:ショウリョウバッタ、オンブバッタ、ショウリョウバッタモドキ
に分けられました。

キリギリス亜目は夜行性、バッタ亜目は昼行性の傾向があり、私たちが暗闇で手を伸ばして物にぶつからないよう歩くように、キリギリス亜目は暗闇での生活に適応して触角が長くなったのかもしれません。
さらに体の構造を観察すると、触角の長さの他にも両亜目の面白い相違点が見つかります。それは「耳(聴覚器官)」の位置です。
「え?昆虫に耳があるの?」と思った人がいるかもしれません。
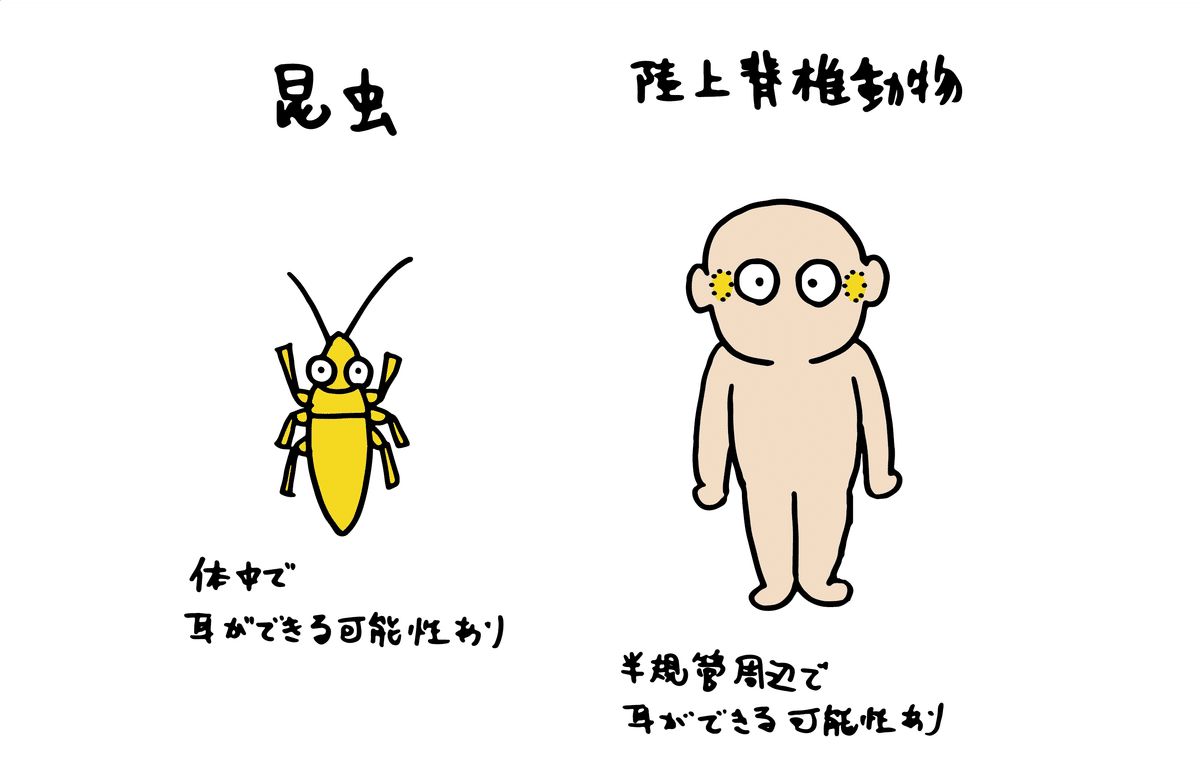
分類学者によって見解が異なることもありますが、昆虫類は27目以上に分けられています。その中で少なくとも7目(バッタ目、カマキリ目、カメムシ目、アミメカゲロウ目、コウチュウ目、チョウ目)から音を感知する耳(聴覚器官)が確認されています。
耳の位置は分類群によって様々で、頭部、胸部、腹部など、細かく分けると少なくとも15の部位で確認されています。
耳の存在意義も、鳴き声を使った個体間のコミュニケーションのために用いたり、コウモリなどの外敵の存在を感知するために用いたり、分類群によって様々です。

耳の位置はというと…
キリギリス亜目は前脚の脛節に、バッタ亜目は胸部にあります。

耳が頭以外にあるなんて、私たち哺乳類からすると奇想天外に感じます。
キリギリス亜目はまるで●クタースランプアラレちゃんの●コチャン大王のようですね(世代によってはネタが分からないかも(^^;))。

同じ目に属するにもかかわらず、キリギリス亜目とバッタ亜目で耳の位置は大きく異なります。このことは、体に耳ができるという進化が私たちヒトよりも簡単に起こることを想像させます。
この耳の位置の違いは、彼らの生活にどの様な影響を与えるのでしょうか?
なぜ昆虫は体の様々な部分に耳を作ることができたのでしょうか?
キリギリスの耳の仕組みはヒトとそっくり!
疑問を解く前に、少しだけ耳から脱線して鼻の話をさせてください。
バッタ目たちの胴体を見てみると、各節に穴があることが分かります。これこそ昆虫が呼吸をするための穴、つまり私たちにとって鼻の機能を持つ部位です。この穴は基本的に胸部の一部と腹部の各節に存在し、「気門」と呼ばれます。
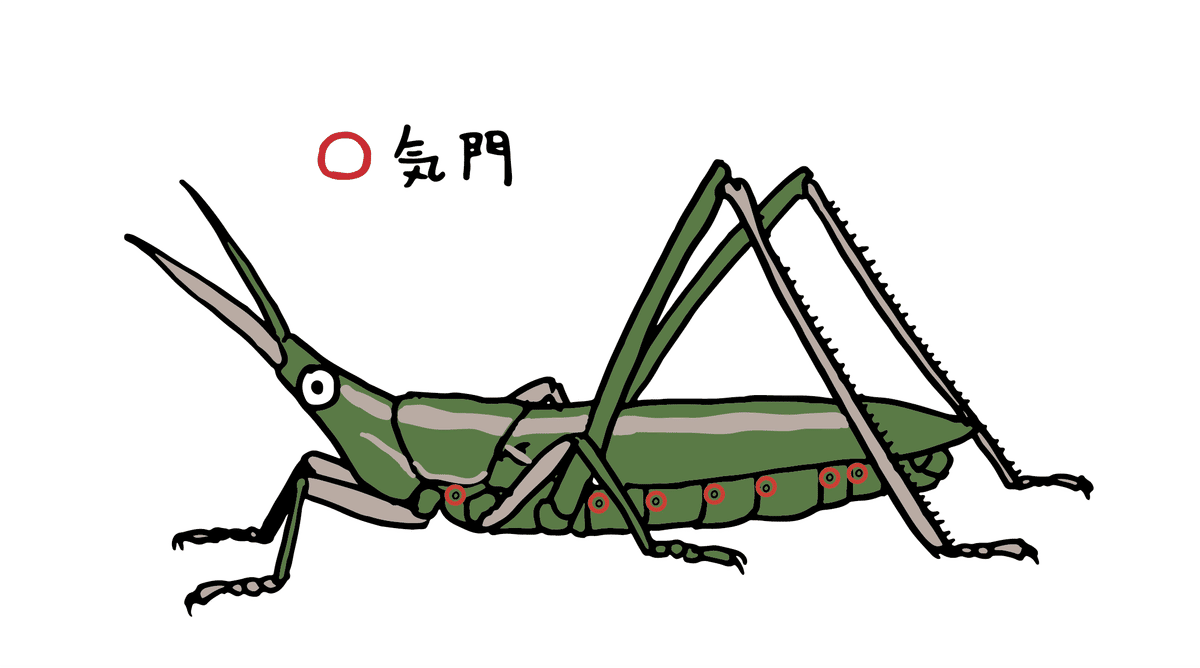
各節にある気門は体の中で繋がっていて、この空気の通り道を「気管」と呼びます。呼吸(大気中の酸素を取り込み、体内の二酸化炭素を出す)を、体中に張り巡らされた気管で行っているのです。
耳の位置然り、肺だけで呼吸している私たち哺乳類とは体の造りが根本的に違うのだと思い知らされますね。
さて、話を耳に戻しますと、先に述べたとおりキリギリス亜目は鳴き声で配偶者を探します。「同種の鳴き声を聞きとる」「どこで鳴いているか捉える」の2つを満たすことが、次世代に自身の遺伝子を残すための鍵となります。
キリギリス亜目のフタホシコオロギ、エンマコオロギを用いた形態学的な研究は、彼らの耳が人間の耳と同じような仕組みで音程を聞きとることを明らかにしました。
コオロギたちの前脚の鼓膜付近にはリンパ液という液体で満たされた部屋があり、そこに音の刺激(振動)を感じとる感覚細胞が並んでいます。前脚の鼓膜から入った音(空気の振動)は気管を伝ってリンパ液の入った部屋に触れている突起を振動させ、部屋を揺らします。部屋の入口では高い音を、末端には低い音を感知する感覚細胞がそれぞれ存在します。
音の性質上、液体では低い音の方が遠くまで振動が伝わりやすく、コオロギの耳はこの性質を上手く利用しているのです。
ヒトの耳でも鼓膜から入った振動を、耳小骨(じしょうこつ)が渦巻管(うずまきかん)へと伝えます。
コオロギの耳の構造と同様に渦巻管の中はリンパ液で満たされています。リンパ液の振動を、渦巻管内の感覚毛が感知し、脳で音情報に変換されます。ヒトの場合でも渦巻管の手前では高い音を、奥の方で低い音を感知します。

音という空気の振動を鼓膜でキャッチして、固体(コオロギでは突起、ヒトの場合は耳小骨)経由で液体(リンパ液)に伝え、感知する。
「一定の音程を聞きとる」というミッションをクリアするために、コオロギとヒトという全く異なる生き物間で同じ仕組みを採用しました。このように異なる分類群間で類似した特徴を持つことを、「収斂(しゅうれん)進化」と呼びます。
同じ物理法則が成り立つ陸上環境で、同じ仕組みの耳という境地にたどり着いたのですね。そんなことを考えながら道端のコオロギを見てみると、何だか同志のような親近感を覚えます。

キリギリスは「ゆっくり」音を聞く
ヒトと同じ仕組みの耳によって、コオロギを含むキリギリス亜目は幅広い音域を感じとり「同種の鳴き声を聞き分ける」ができているようです。
では、どのように「どこで鳴いているか捉える」を可能にしているのでしょうか?
そもそも私たちは無意識のうちに音源(音の発生位置)を感じとることができます。
これは耳の位置の左右差を利用しているためです。
空気の振動である音が正面で起こった場合、振動は左右の耳に同時に届きます。右方向で起こった場合、空気の振動はまず右の耳に届き、次に頭を回り込んで左の耳に届きます。空気を介して左右の耳が振動をキャッチするまでのタイムラグ(時間差)によって音源を特定できるのです。
私たちヒトが音源を特定する過程を科学館の音ゾーン「どう聞こえる?」という展示で学ぶことができます。
キリギリス亜目にも、音源を特定するための工夫が2つあります。
1つ目は、ヒトと同じように左右の耳の位置をできるだけ離すこと。
他の昆虫の耳が頭部、胸部、腹部など体幹にある一方で、キリギリス亜目は前脚の端に存在します。前脚を広げることで、左右の耳の位置をできるだけ離し、左右の音が到達するまでのタイムラグを生み出すことに繋がっていると考えられます。

しかし、所詮は昆虫の体です。ヒトの頭と比べると左右の耳の距離はとても小さく、さらなる工夫が必要です。
もう一つの工夫点は、音を「ゆっくり」聞くことです。
キリギリス亜目の一種は前脚の付け根の気門から空気の振動を取り入れることが知られています(ここでは音響気門と呼びます)。音響気門から入った空気の振動は、前脚の気管をとおって、脚の関節を順に進み、最終的に耳に伝わります。
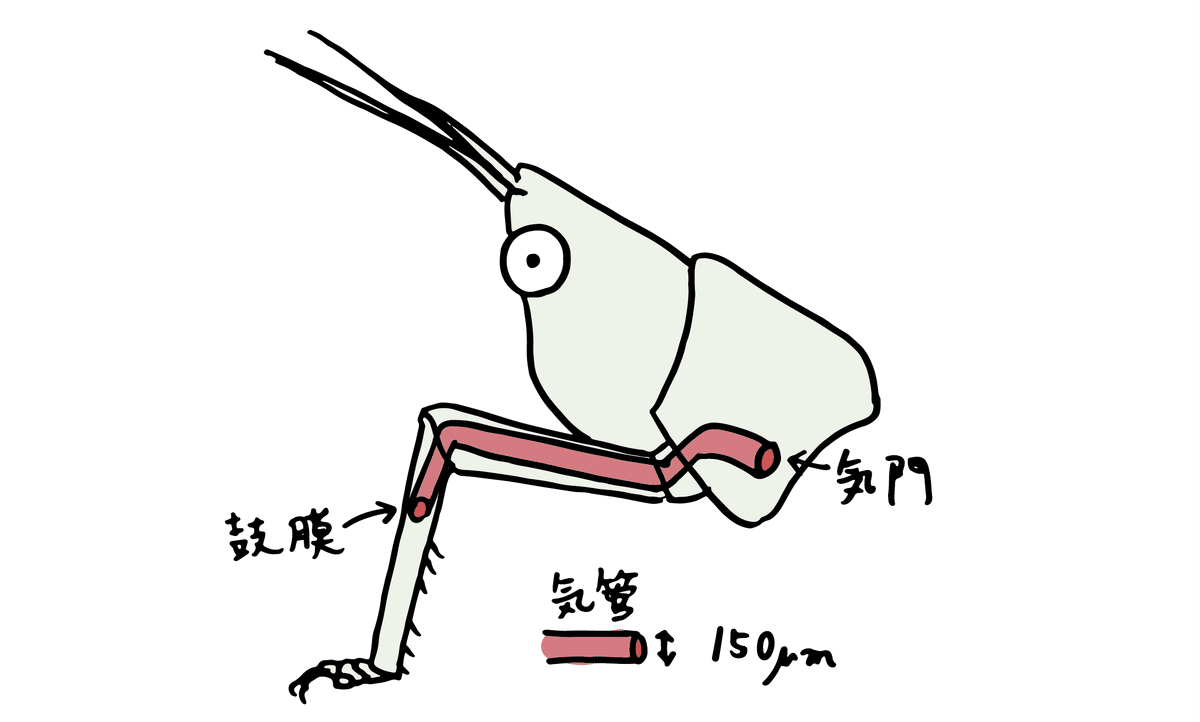
空気の振動が平均直径150マイクロメートルという狭い気管を進むにつれて音は「減速」します。
また、気管では呼吸も同時に行なわれているため大気中よりも二酸化炭素濃度が高く、この空気の組成の違いも音の減速に拍車をかけます。実験では、大気中を伝わる音よりも、最大で秒速100 m程度減速することが示されています。
音をゆっくり聞くことができれば、音が聴覚器官に届く際の左右間のタイムラグも大きくなります。
以上の耳の位置と、気管を利用した音の減速によって、キリギリス亜目たちは音源の位置を正確に把握することができると考えられています。
ここまで耳の仕組みについて「キリギリス亜目」とひとくくりにしてきましたが、全てのキリギリス亜目に同じような耳の仕組みがあるわけではありません。また、翅の退化した分類群、例えばカマドウマ科やコロギス科などは鳴くことがなく、耳も退化しています。
逆に言えば、耳を持つほとんどの種は音を使ったコミュニケーションをとることから、キリギリス亜目の耳と発音器官は密接な関係にあると考えられています。
昆虫と哺乳類の耳の歴史を紐解くと…
バッタ亜目では、耳があっても音を使ったコミュニケーションをとらないことが多々あり、耳と発音器官の関係性はキリギリス亜目ほど強くありません。
キリギリス亜目とバッタ亜目の生態・進化の違いは、脚の先端に耳を作ったか体幹に耳を作ったかという耳の位置の違いによるものかもしれません。
この様に、昆虫の世界では、耳は同じグループの中でも複数回進化して登場したり退化したりする比較的「お手軽な」感覚器官なことが分かります。
コオロギの音を感じとる感覚細胞はリンパ液を介した特別なものでしたが、多くの昆虫の場合、造りがより簡易的な「機械受容器」と呼ばれる器官で成り立っています。これはもともとクチクラという薄いタンパク質でできた外骨格のひずみを感知する部分が変化したものです。
簡単に整理すると、昆虫が音を聞く仕組みは、①機械受容器、②膜で捉えた振動を体内に伝える気管、③外部の空気の振動を捉える薄い膜(鼓膜、気門、毛など)、という3つから成り立っています。
これら3つは、どの昆虫にも全身にまんべんなく存在します。昆虫の耳の進化が起こりやすい事も納得できますね。
昆虫の耳の歴史をさかのぼると、3億1千万年前(石炭紀)の地層から発音機能を有する翅をもつティタノプテラ目の一種の化石が発掘されています。もちろん発音することが音を用いたコミュニケーションをしていた証拠、つまり耳をもつことにはなりません。
しかし、約2億年前には現在のバッタ目のような前脚に鼓膜を持つ昆虫が現れ始めました。同じく現代の昆虫の祖先種たちの分化が完了し、この時代に耳の仕組みをもつ分類群が複数いた可能性があります。
私たちヒトの耳の進化過程もみてみましょう。
陸上の脊椎動物の場合、昆虫の①機械受容器の役割は、「半規管(はんきかん)」と呼ばれる平衡(へいこう)感覚や加速度を感知するための器官がもとになっています(現在の渦巻管)。
半規管は5億年以上前に当時水中生活していた陸上脊椎動物の祖先が既に持っていました。液体の振動を外部から体内へ直接伝え、半規管を用いて水中の低周波を聞きとっていたと考えられています。
しかし、上記の水中用の聴覚器官は、陸上では上手く機能しませんでした。
陸上で耳を機能させるためには、液体ではなく大気の振動をキャッチするための鼓膜、そしてその振動を半規管へ伝える骨(現在の耳小骨)が必要だったのです。
鼓膜については化石に残らないため詳しい歴史的変遷は分かっていません。
一方で鼓膜の振動を伝える骨は、顎を支える骨の一部が起源であることが化石情報から分かっています。4億年前には、アカントステガ、イクチオステガなどの四足歩行動物の顎の骨の一部が、耳として機能していたのではと推察されています。
時代は進み、2億年前に誕生した小型哺乳類の半規管は、約2㎜と小さく、形も直線的で単純でした。しかし、ジュラ紀(1億4500万年以前)に生息していた哺乳類の一種の半規管は、3/4ほど渦巻いていました。もう渦巻管と呼んでも差し支えないかもしれませんね。
渦巻管の長さは聴力の周波数範囲と相関しているため、この時代の哺乳類は高音域もある程度聞きとれていたと考えられます。
陸上脊椎動物の分類群間、例えば鳥類と哺乳類では耳小骨の数が異なる事、また発生学的な研究によって鼓膜の形成過程が異なり、それぞれ個別に鼓膜を獲得したことが分かっています。
しかし、半規管・耳小骨・鼓膜という3点セットの基本構造は陸上脊椎動物で共通しており、この基本構造から外れた進化(例えば、キリギリス亜目や●コチャン大王のように足に耳ができる)は起こりませんでした。
おわりに
秋の虫から、キリギリス亜目とバッタ亜目の違い、キリギリス亜目の驚くべき耳の機能、昆虫と哺乳類の進化の道のり、とかなり盛りだくさんな内容をお届けしました。
とは言っても、今回ご紹介した耳に関する情報はごく一部であること、また現在も多くの研究がなされ、新知見によって情報は日々更新されていることをお断りいたします。
身近な秋の虫を見つめ直すことで、私たちヒトと昆虫の体の仕組みは全く異なることが分かりました。
タンパク質でできた外骨格をもつ昆虫と、頭部の半規管に頼った耳を作り続ける陸上脊椎動物。それぞれの身体的な基盤の違いによって、全く異なる耳の進化が起こりました。
そんなことを考えると、秋の虫の声をいつもと違った気持ちで聞くことができるかもしれません。残り少ない秋を大切に感じてみましょう。

参考資料
Celiker, E., Jonsson, T. & Montealegre-Z, F. The Auditory Mechanics of the Outer Ear of the Bush Cricket: A Numerical Approach. Biophys. J. 118, 464–475 (2020).
西野浩史. 昆虫の聴覚器官 ―その進化―. 比較生理生化学 23, 26–37 (2006).
Nishino, H., Domae, M., Takanashi, T. & Okajima, T. Cricket tympanal organ revisited: morphology, development and possible functions of the adult-specific chitin core beneath the anterior tympanal membrane. Cell Tissue Res. 2019 3772 377, 193–214 (2019).
Schubnel, T. et al. Sound vs. light: wing-based communication in Carboniferous insects. Commun. Biol. 2021 41 4, 1–11 (2021).
Song, H. et al. Phylogenomic analysis sheds light on the evolutionary pathways towards acoustic communication in Orthoptera. Nat. Commun. 2020 111 11, 1–16 (2020).
Veitch, D. et al. A narrow ear canal reduces sound velocity to create additional acoustic inputs in a microscale insect ear. Proc. Natl. Acad. Sci. 118, (2021).
Warren, B. & Nowotny, M. Bridging the Gap Between Mammal and Insect Ears – A Comparative and Evolutionary View of Sound-Reception. Front. Ecol. Evol. 0, 504 (2021).

いいなと思ったら応援しよう!
